This is a short introduction to my research form the last couple of years. Please also have a look at a talk that I prepared for the Topical Problems of Biophotonics meeting in July 2019. Download (40 MB)
For more details I would like to point you to our publications.
Event Driven Acquisition
While many biological events are highly dynamic, microscopes are usually preset to perform a fixed imaging sequence. We developed an event-driven acquisition framework, in which neural-network-based recognition of specific biological events triggers real-time control in an instant structured illumination microscope. This allows us to adjust the imaging to the dynamic nature of the sample and capture more details of the biological events of interest.

Molecular Motors
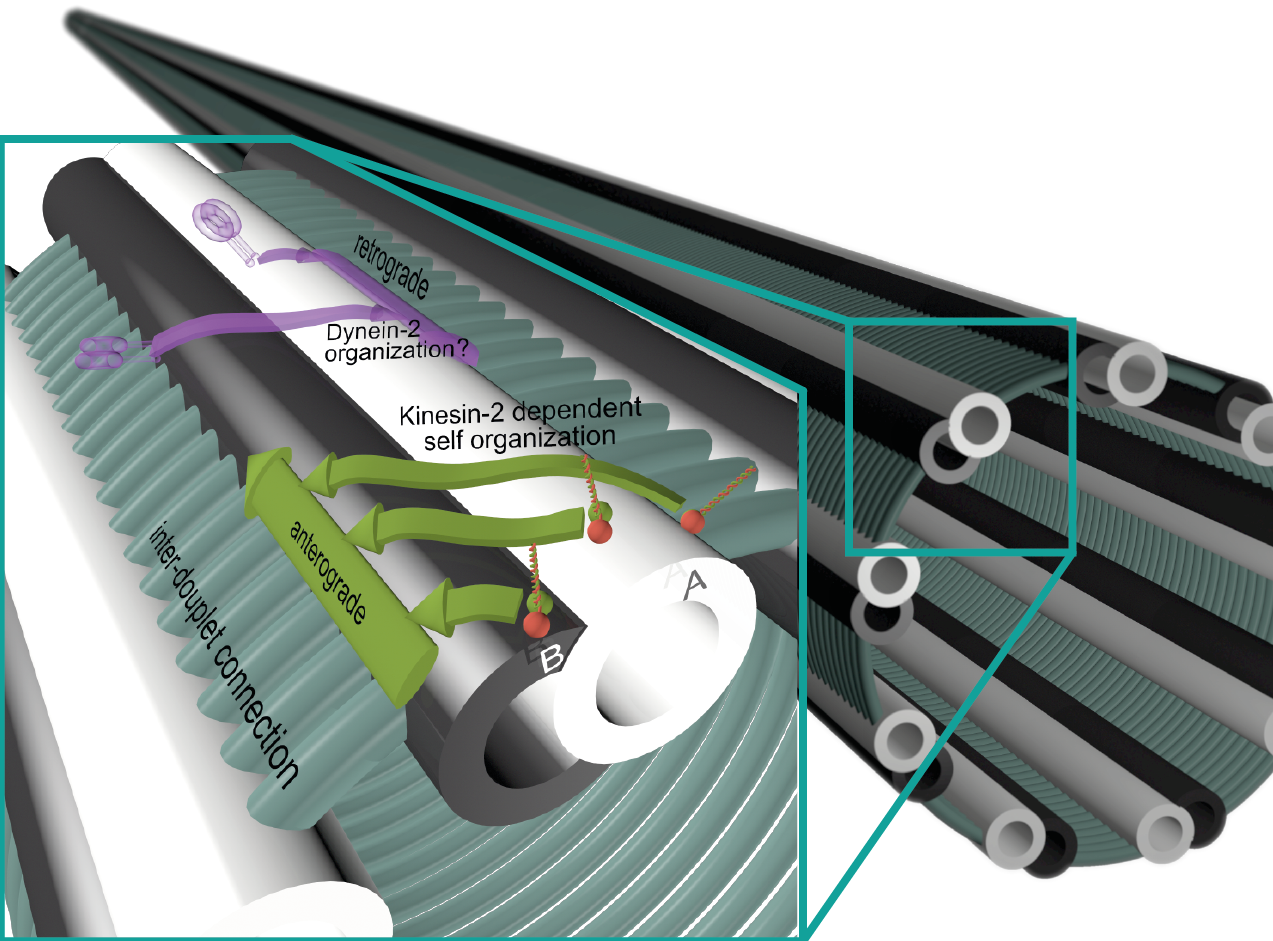
Molecular motors transport cargo in cells that can not be distributed effectively via diffusion. Using ATP as a fuel, they can achieve remarkable performance for a motor. Considering they are eight orders of magnitude smaller than we are, they walk at a normalized speed of 144 km/h. Kinesins walk on filaments called microtubules, or in the special environment of a cilium on a axoneme. The motors support the build-up and maintenance of the structure of the cilium in a process called intraflagellar transport (IFT).
Intraflagellar Transport
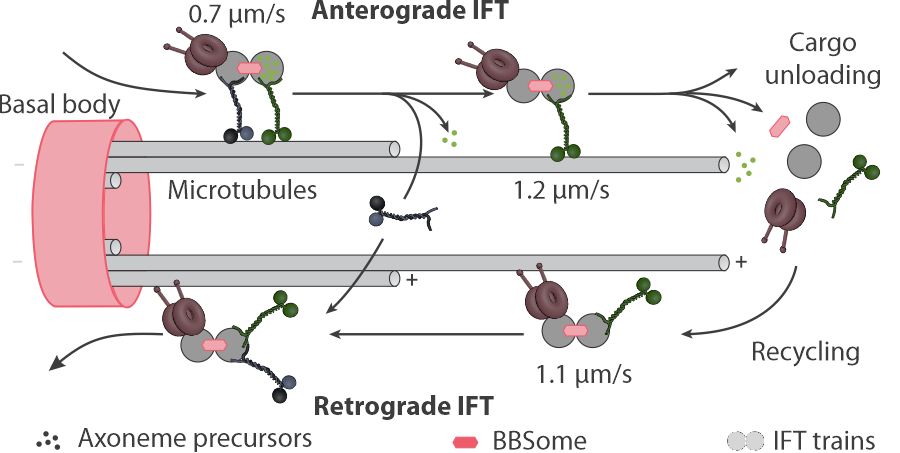
adjusted (Scholey J.M., 2013, Annu Rev Cell Dev Biol)
Cilia are used for locomotion or sensory purposes in virtually all eukaryotes. This figure depicts the process in c. elegans. Two kinesin-2 motors transport the IFT cargo (IFT trains) to the tip. The cargo is made up from repeating protein complexes that consist of several 10th of proteins each, including a deactivated dynein-2 motor. At the tip, the cargo is remodeled and used to build the axoneme. The scaffold of the IFT train is transported back to the base of the cilium by the dynein-2. The cargo now also includes the inactivated kinesin-2 motors as a cargo.
Most kinesin-2 motors are heterodimers, meaning that they are made up of two different subunits. Their activity on the special filament axoneme and the special property of them being heterodimers has sparked important question about their walking mechanism. The walking mechanism of molecular motors is best studied with a technique called FIONA. During my Masters' Thesis, I built a microscope that is optimized for this super-resolution technique.
mcFIONA setup

adjusted (Scholey J.M., 2013, Annu Rev Cell Dev Biol)
Cilia are used for locomotion or sensory purposes in virtually all eukaryotes. This figure depicts the process in c. elegans. Two kinesin-2 motors transport the IFT cargo (IFT trains) to the tip. The cargo is made up from repeating protein complexes that consist of several 10th of proteins each, including a deactivated dynein-2 motor. At the tip, the cargo is remodeled and used to build the axoneme. The scaffold of the IFT train is transported back to the base of the cilium by the dynein-2. The cargo now also includes the inactivated kinesin-2 motors as a cargo.
Most kinesin-2 motors are heterodimers, meaning that they are made up of two different subunits. Their activity on the special filament axoneme and the special property of them being heterodimers has sparked important question about their walking mechanism. The walking mechanism of molecular motors is best studied with a technique called FIONA. During my Masters' Thesis, I built a microscope that is optimized for this super-resolution technique.
mcFIONA: multi-color Fluorescence Imaging with One Nanometer Accuracy
This is the design of the super-resolution microscope that I built during 2016 and 2017. It is optimized for stepping detection of molecular motors with total internal reflection fluorescence (TIRF) illumination. With this setup I was able to study the stepping mechanism of kinesin-2 in great detail. The basic stepping mechanism of a kinesin-2 is shown below in a simplified form.
The kinesin walking mechanism

Using a motor that is labeled on both heads, I can follow every step of the motor and extract the basic stepping parameters. Interesting for the determination of the stepping behavior is the step size, the distance between to binding events of a head. Also the timing of the steps is characterized by the dwell times. The time a motor spends attached to the filament between steps.
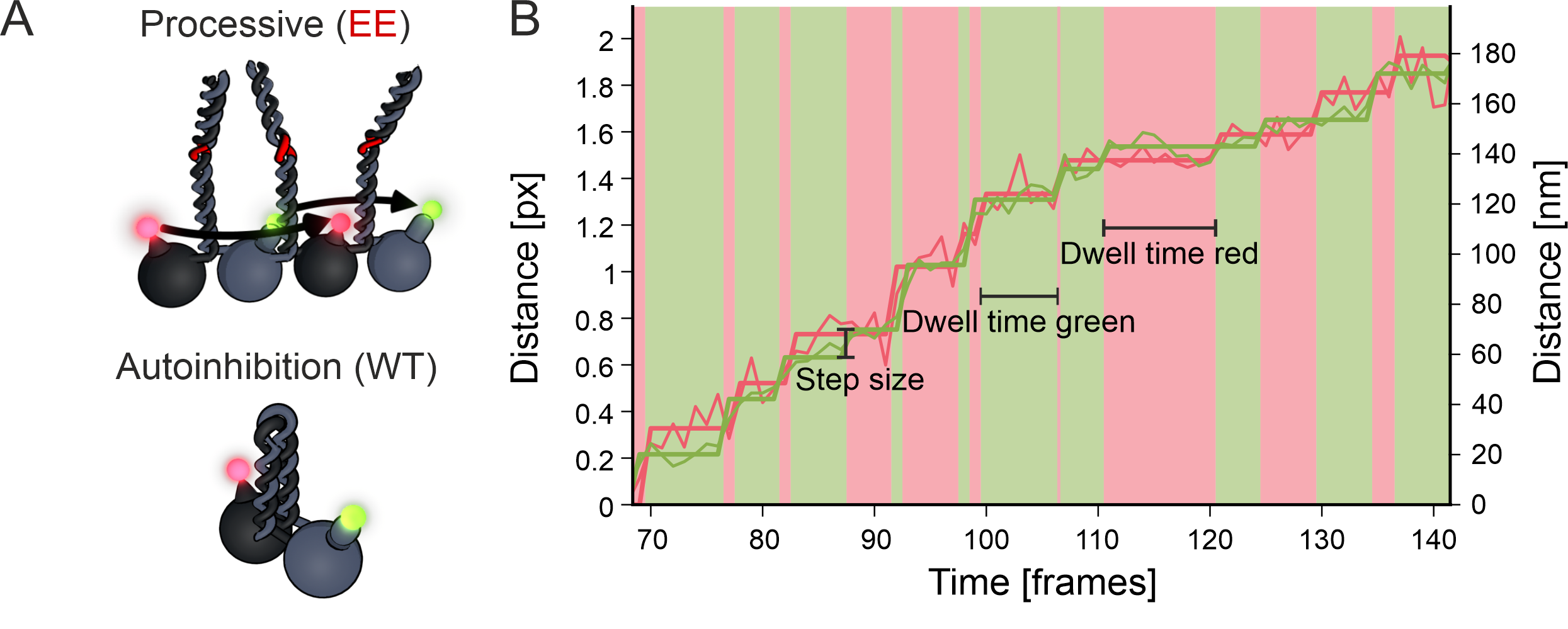
Kinesin-2 limping
By measuring these parameters for both heads on one motor at the same time, I was able to show, that the different heads in the heterodimeric motor actually show different stepping behavior. The distribution of dwell times of steps for one subunit (KLP20) is close to a single exponential, while the other distribution is close to a convolution of two exponentials.
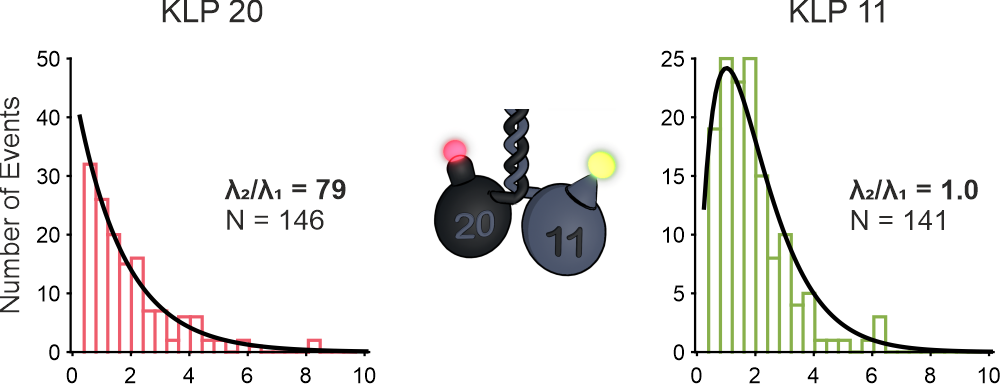
Kinesin-2 differentiates between microtubules and axonemes
As kinesin-2 is mainly active in cilia, it walks on its special filament called axoneme. I measured the step sizes for a kinesin-2 motor on both axonemes and microtubules. On microtubules, the step size is ~13 nm, while it is 16~nm on the axoneme. This reflects a property of kinesin-2 motors to step sideways on the microtubule. This can also be seen in the movement of the motors, as the traces are straight on axonemes, but reflect a sinusoid on microtubules.
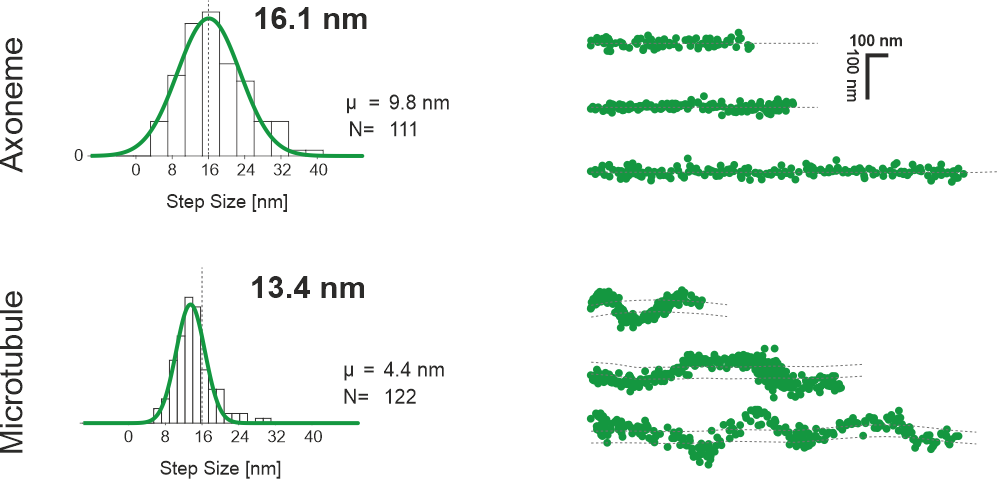
This shows, that kinesin-2 motors walk straight on axonemes, while they spiral around a microtubule. For the IFT, this means that the motors spiral until they reach the axoneme-specific barrier between microtubule doublets. This directs the traffic to one side of the microtubule doublet, freeing the other side for the opposite traffic coming to the base. This ensures optimal IFT in the dense environment of the cilium.